
How do you determine whether or not a fossil belongs to the human lineage?
It’s important to distinguish between two things: Hominins and the human line. Hominins include representatives of the Homo genus and all fossils closer to us than chimpanzees. The human lineage refers to the ancestors of our species, Homo sapiens. So, among the Hominins, paleoanthropologists are looking for those that could be at the origin of the human lineage. To do this, they defined criteria such as mode of locomotion.
Indeed, although some anatomical criteria exist, it is mainly bipedal characteristics that are used to determine whether or not a fossil belongs to the human lineage. Why bipedalism? Quite simply because it’s the preferred mode of locomotion for our species! We must be careful, however, as Homo sapiens is not the only species capable of bipedalism. In fact, the ability to move in a bipedal manner, i.e. on both lower limbs, appeared well before the emergence of the human lineage around 7 million years ago (later abbreviated to Ma). Indeed, this type of locomotion appeared as early as 250-200 Ma. It is found, for example, in certain dinosaurs. Even today, other animals such as birds and kangaroos move around the ground in a bipedal fashion. Among primates, the order to which Homo sapiens belongs, many species are capable of bipedalism. In fact, our species is the only primate to practice a single mode of locomotion for which our bodies are hyper-specialized. Furthermore, scientists agree to no longer speak of a single biped, but of bipeds, to emphasize that different bipedal locomotions have existed within the Hominins. To find out more about the skeletal adaptations required for bipedalism, click here.
Exclusive bipedalism therefore seems to be a feature of our lineage. This is probably why studies on the bipedalism of early Hominins are at the heart of the debate. What about our three candidates?
Sahelanthropus tchadensis, one of the oldest candidates for the origin of the human lineage
This species was discovered between 2001 and 2002 by a Franco-Chadian team at Toros-Menalla in Chad (Africa). Several cranial remains (3 mandibles and several isolated teeth), belonging to at least 3 different individuals, as well as an almost complete skull were unearthed. What’s more, this skull is nicknamed Toumaï! A few postcranial remains (a left femoral shaft, two right ulnas and a left ulna) were also discovered. The only problem with the latter remains…they were not discovered in direct connection with the cranio-dental remains. Consequently, it is impossible to state with certainty that they belong to S. tchadensis! Nevertheless, the discoverers of the fossils feel that the most parsimonious hypothesis is to consider that these postcranial remains belong to the only species present on the site, S. tchadensis.
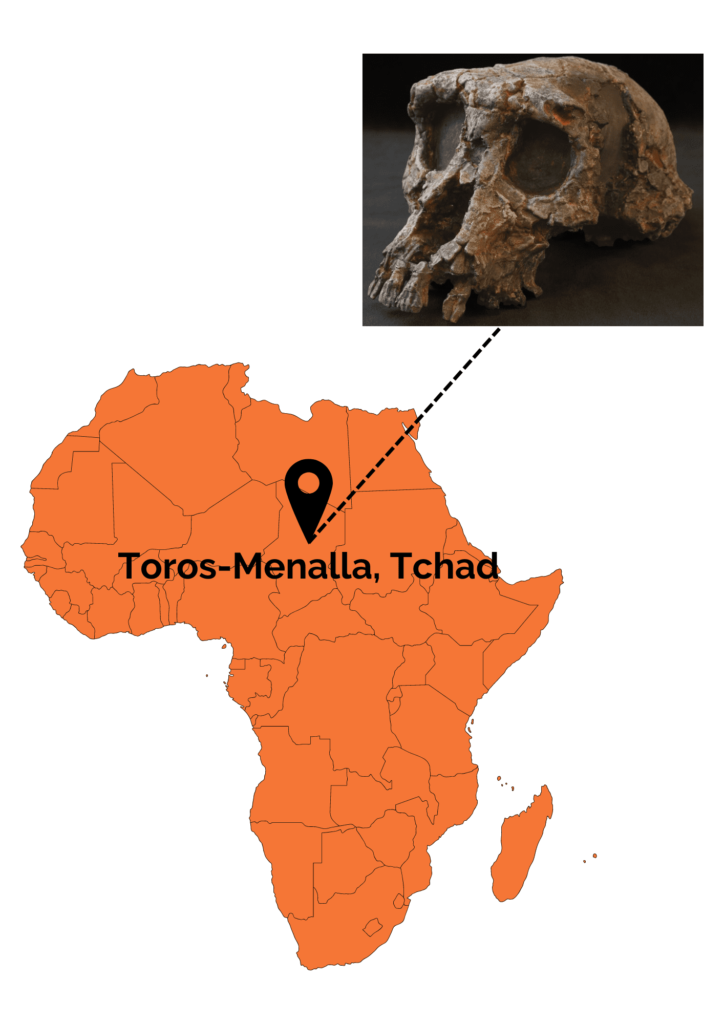
The fossil remains of S. tchadensis have been dated to between 7 and 6 Ma using two methods. The first is biochronology, i.e. a chronological estimate based on the degree of evolution of fossil faunal remains found within the same stratigraphic zone. The second method used is cosmogenic nuclide dating (Aluminum 26Al/Beryllium10Be). According to the fauna associated with the fossil remains and the palynological studies carried out in the region, S. tchadensis lived in an environment combining forests, savannahs, grasslands and bodies of water. This region is now desert.
Several anatomical features bring us closer together S. tchadensis of later Hominins, such as small canines, absence of diastema (space between incisors and canines due to the large size of the latter) and a more anteriorly positioned foramen magnum than in non-human great apes. These characteristics distinguish S. tchadensis from gorilla ancestors, a hypothesis put forward at the time the fossils were discovered. Nevertheless, the cranial features must be interpreted with caution, as the skull is deformed by the weight of the sediment. As a result, certain characters have been deduced from virtual reconstructions. Other features, such as a small cranial capacity of around 350 cm3, bring S. tchadensis closer to non-human primates.

Toumaï is far from being unanimously accepted by the scientific community. For its discoverers, the anterior position of the foramen magnum indicates bipedal locomotion. For many other scientists, however, this alone is not enough, especially as the base of the skull, where the foramen magnum is located, is severely damaged. What’s more, the few postcranial remains found, notably the femurs, do not seem to reveal any clear adaptations to bipedalism. The debate continues with a focus on dental residuals. Apart from bipedalism, the size of the canines (reduced in H. sapiens) is also a criterion used to assign a fossil to the Hominins. S. tchadensis has a priori small canines, but only if we consider that the skull found belongs to a male. This deduction is based solely on the size and width of the supraorbital torus. However, the link between supraorbital torus size and gender is far from unanimous, as it is considered an unreliable trait. For some researchers, the most prudent hypothesis would therefore be to consider that the skull belongs to a female. The small size of the canines would therefore not be the result of any link with Hominins, but simply linked to a phenomenon of sexual dimorphism.
The saga continues with a new article published in 2022 in Nature where a femur, discovered more than 20 years ago, has just been officially described and whose study shows that a dozen characters demonstrate the bipedal ability of S. tchadensis although he was certainly still climbing trees. Finally, in 2023, Marc R. Meyer et al. published a study that raised the hypothesis of knuckle-walking locomotion! This would exclude S. tchadensis from the Hominins.
As you can see, although still considered a potential ancestor of the human lineage, the position of S. tchadensis is hotly debated.
Orrorin tugenensis
The species Orrorin tugenensis was created in 2000 following the discovery of a dozen dental and postcranial remains on Tugens Hill in Kenya by Martin Pickford and Brigitte Senut. These remains are dated between 6.1 and 5.7 Ma.
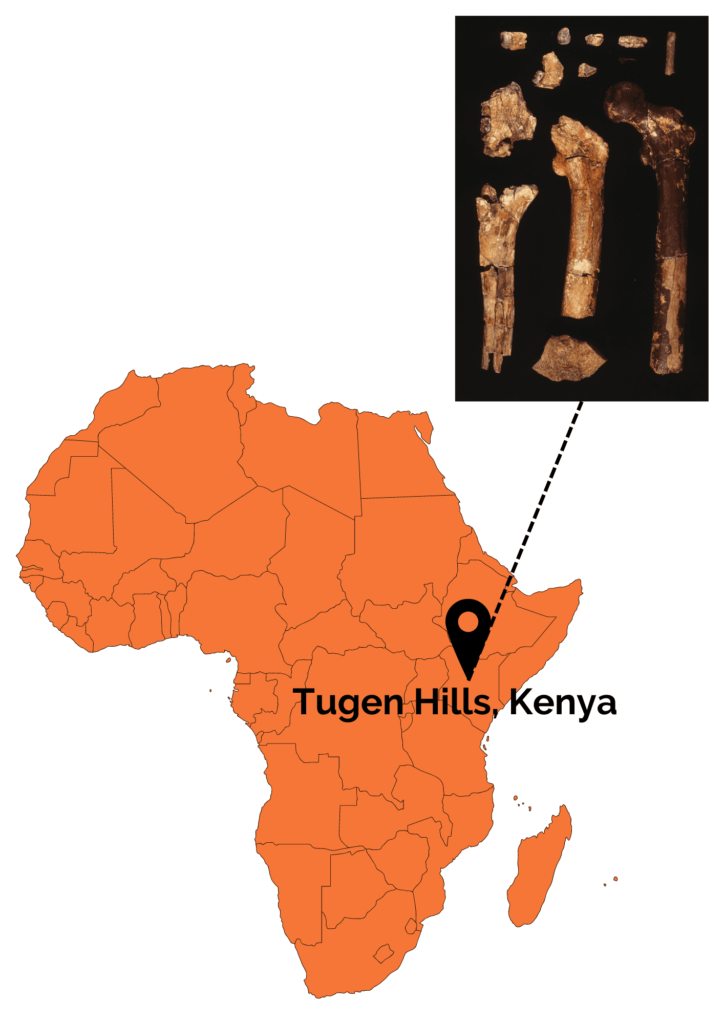
Several morphological features bring O. tugenensis closer to the more recent Hominins. In terms of dentition, the molars found are square-shaped and small compared to other non-human primates. In addition, the canines are reduced in size and there is an absence of diastema. Unlike chimpanzees, Orrorin’s upper molars show different degrees of wear, suggesting differential molar growth. In fact, the first molar, which is the most worn, would have come out before the second molar and finally the third molar, which is hardly worn at all. This differentiated growth of molars is found not only in H. sapiens, but also in Australopithecines.
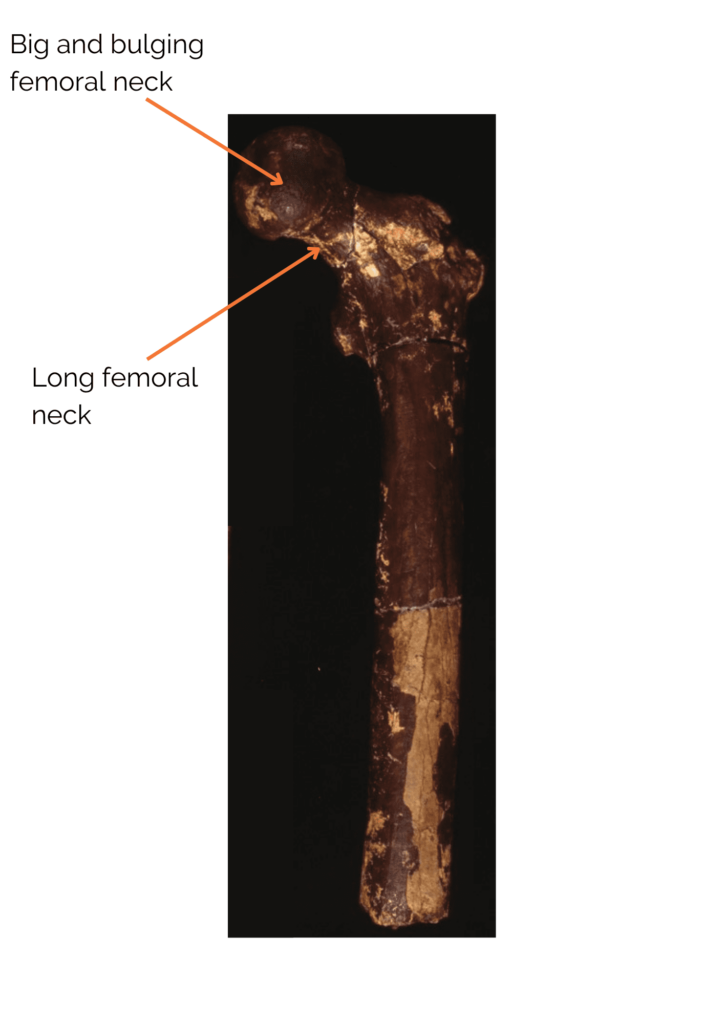
Postcranial remains show several signs of regular bipedal locomotion. These include a long femoral neck with a large, globular head, similar to that of Homo sapiens. In addition, the cortical bone at the femoral neck is very thick. This is a good indication of bipedalism, as it is interpreted in bipeds as reinforcement to support the weight of the trunk without risk of breakage. Nevertheless, although Orrorin was probably an occasional or regular biped, his bipedalism was probably not like ours. It’s also worth noting that the bones of the upper limbs, notably the curved phalanx of the hand, show adaptations to tree climbing. Orrorin was therefore capable of both arboricolia and bipedalism! In terms of its environment, studies have shown that Orrorin evolves in a dry temperate forest environment with the presence of wetlands. This has been much debated within the scientific community, as it contradicts the theory of the emergence of bipedalism through body straightening due to an open environment. Studies of its teeth suggest that it was a frugivore or omnivore.
The Ardipithecus genus
The latest candidate for the title of oldest Hominin is the genus Ardipithecus. Two species belong to this genus:
- Ardipithecus kadabba, dated to between 5.77 and 5.2 Ma, whose fossils have been found in various localities in the middle Awash valley in Ethiopia.
- Ardipithecus ramidus, also discovered in Ethiopia’s Middle Awash Valley and dated at around 4.4 Ma

A. kadabba has many primitive features, such as large canines implying the presence of a diastema, and a particularly strong elbow joint indicating brachiation locomotion. Similarly, the phalanges of the hands and feet are long and curved, indicating a capacity for arboricolia. Nevertheless, there are a few features that bring this species closer to the human lineage. This is the case, for example, of the articular surface of a proximal foot phalanx, which has a dorsal inclination. This trait is considered unique to bipeds, linking A. kadabba to more recent Hominins. A. kadabba is sometimes seen as the ancestor of the second known species belonging to the genus Ardipithecus: Ardipithecus ramidus.
Like its potential ancestor, Ardipithecus ramidus also displays features that indicate both arboreal and bipedal locomotion.
Although the place of these 3 genera within the Hominins remains debated, their importance in understanding the origin of the human lineage is certain! Indeed, these specimens are from a period when our ancestors diverged from the ancestors of chimpanzees, our closest cousins, and thus illustrate the beginning of human history.
Feel free to ask us questions and give us feedback on the blog. You can also contact us by e-mail. You can also follow us on Instagram, Facebook, Twitter, TikTok, LinkedIn and YouTube !
See you soon,
The Prehistory Travel team.
Bibliography :
[1] Michel Brunet, “Sahelanthropus tchadensis dit “Toumaï” : le plus ancien membre connu de notre tribu”, Bulletin de l’Académie Nationale de Médecine, 204 (2020)
[2] Michel Brunet et al, “Toumaï, Miocène supérieur du Tchad, le nouveau doyen du rameau humain”, Comptes Rendus Palevol, 3 (2004).
[3] G. Daver et al, “Postcranial evidence of late Miocene hominin bipedalism in Chad”, Nature, 609 (2022), https://doi.org/10.1038/s41586-022-04901-z
[4] Martin R. Meyer et al, “Knuckle-walking in Sahelanthropus? Locomotor inferences from the ulnae of fossil hominins and other hominoids”, Journal of Human Evolution, 179 (2023), 103355
[5] M. Pickford et al, “Bipedalism in Orrorin tugenensis revealed by its femora”, Compte rendus Palevol, 1 (2002)
[6] Y. Sawada, “The age of Orrorin tugenensis, an early hominid from the Tugen Hills, Kenya”, Comptes rendus Palevol, 1 (2002)
[7] B. Senut, ” Orrorin tugenensis et les origines de l’homme: une synthèse”, Bulletin de l’Académie Nationale de Médecine, 204 (2020)
[8] Peter A. Stamos et al, “Hominin locomotion and evolution in the Late Miocene to Late Pliocene”, Journal of Human Evolution, 178 (2023), 103332
[9] Tim D. White et al, “Ardipithecus ramidus and the Paleobiology of early hominids”, Science, 326 (2009)


