
Comment déterminer si un fossile appartient ou non à la lignée humaine ?
Il est important de distinguer deux choses : les Hominines et la lignée humaine. Les Hominines comprennent les représentants du genre Homo et tous les fossiles plus proches de nous que des chimpanzés. La lignée humaine, quant à elle, désigne les ancêtres de notre espèce, Homo sapiens. Ainsi, parmi les Hominines, les paléoanthropologues cherchent ceux pouvant être à l’origine de la lignée humaine. Pour cela, ils ont défini des critères comme le mode de locomotion.
En effet, bien que quelques critères anatomiques existent, ce sont surtout des caractères liés à la bipédie qui sont utilisés pour déterminer si un fossile appartient ou non à la lignée humaine. Pourquoi la bipédie ? Tout simplement car il s’agit du mode de locomotion privilégié que notre espèce utilise ! Il faut néanmoins faire attention car Homo sapiens n’est pas la seule espèce capable de bipédie. En réalité, la capacité à se déplacer de façon bipède, c’est-à-dire sur les deux membres inférieurs, est apparue bien avant l’émergence de la lignée humaine vers 7 millions d’années (abrégé par Ma par la suite). En effet, dès 250-200 Ma, ce type de locomotion apparaît. On le retrouve par exemple chez certains dinosaures. Encore aujourd’hui, d’autres animaux comme les oiseaux ou les kangourous se déplacent au sol de façon bipède. Parmi les primates, ordre auquel appartient Homo sapiens, de nombreuses espèces sont capables de pratiquer la bipédie. En réalité, notre espèce est la seule parmi les primates à pratiquer un seul mode de locomotion pour lequel notre corps est hyper spécialisé. Par ailleurs, les scientifiques s’accordent à ne plus parler d’une seule bipédie mais des bipédies pour souligner que différentes locomotions bipèdes ont existé au sein des Hominines. Si vous souhaitez en savoir plus sur les adaptations squelettiques nécessaires à la bipédie, cliquez sur ce lien.
La bipédie exclusive semble donc être une caractéristique de notre lignée. C’est probablement pour cela que les études sur la bipédie des premiers Hominines sont au cœur des débats. Qu’en est-il de nos trois candidats ?
Sahelanthropus tchadensis, l’un des plus anciens candidats à l’origine de la lignée humaine
Cette espèce est découverte entre 2001 et 2002 par une équipe franco-tchadienne à Toros-Menalla au Tchad (Afrique). Plusieurs restes crâniens (3 mandibules et plusieurs dents isolées), appartenant au moins à 3 individus différents, ainsi qu’un crâne quasiment complet sont mis au jour. Par ailleurs, ce crâne est surnommé Toumaï ! Quelques restes postcrâniens (une diaphyse fémorale gauche, deux ulnas droits et un ulna gauche) sont également découverts. Seul problème à ces derniers restes…ils n’ont pas été découverts en lien direct avec les restes cranio-dentaires. Par conséquent, il est impossible d’affirmer avec certitude que ces derniers appartiennent bien à S. tchadensis ! Néanmoins, les découvreurs des fossiles ont estimé que l’hypothèse la plus parcimonieuse est de considérer que ces restes postcrâniens appartiennent à la seule espèce présente sur le site, S. tchadensis.
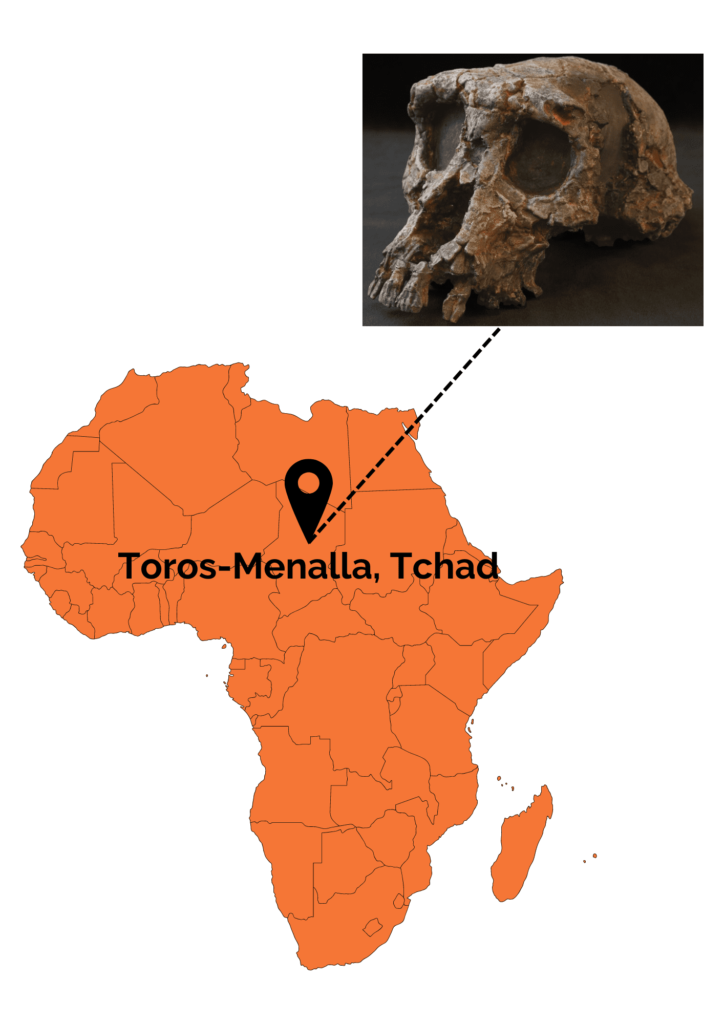
Les restes fossiles de S. tchadensis ont été datés entre 7 et 6 Ma grâce à deux méthodes. La première est la biochronologie, c’est-à-dire une estimation chronologique à partir du degré d’évolution des restes fossiles de faune retrouvés au sein d’une même zone stratigraphique. La seconde méthode employée est une méthode de datation par les nucléides cosmogéniques (Aluminium 26Al/Béryllium 10Be). Toujours d’après la faune associée aux restes fossiles et aux études palynologiques effectuées dans la région, S. tchadensis aurait vécu dans un environnement mêlant forêts, savanes, prairies et étendues d’eau. Cette région est aujourd’hui désertique.
Plusieurs caractéristiques anatomiques rapprochent S. tchadensis des Hominines plus tardifs tel que des canines de petite taille, une absence de diastème (espace entre les incisives et les canines dû à la grande taille de ces dernières) ainsi qu’un trou occipital en position antérieure plus antérieure que chez les grands singes non humains. Ces caractéristiques distinguent S. tchadensis des ancêtres des gorilles, une hypothèse avancée au moment de la découverte des fossiles. Néanmoins, les caractères crâniens sont à interpréter avec précaution car le crâne est déformé par le poids des sédiments. De ce fait, certains caractères ont été déduits à partir de reconstructions virtuelles. D’autres caractères, comme une faible capacité crânienne d’environ 350 cm3, rapprochent S. tchadensis des primates non-humains.

Toumaï est loin de faire l’unanimité au sein de la communauté scientifique. Pour ses découvreurs, la position antérieure du trou occipital indique une locomotion bipède. Néanmoins, pour de nombreux autres scientifiques, ce caractère seul n’est pas suffisant et ce, d’autant plus que la base du crâne, l’endroit où se situe le trou occipital, est fortement endommagée. De plus, les quelques restes postcrâniens retrouvés, notamment les fémurs, ne semblent pas révéler de claires adaptations à la bipédie. Le débat continue si l’on s’intéresse cette fois-ci aux restes dentaires. Hormis la bipédie, la taille des canines (réduites chez H. sapiens) est un critère également utilisé pour rattacher un fossile aux Hominines. S. tchadensis présente a priori des canines de petite taille mais seulement si l’on considère que le crâne retrouvé appartient à un mâle. Cette déduction est réalisée seulement sur la taille et la largeur importante du torus supraorbitaire. Or, le lien entre taille du torus supraorbitaire et sexe est loin de faire l’unanimité car il s’agit d’un caractère considéré comme peu fiable. Pour certains chercheurs, l’hypothèse la plus prudente serait donc de considérer que le crâne appartient à une femelle. La petite taille des canines ne serait donc pas le résultat d’un quelconque lien avec les Hominines mais tout simplement lié à un phénomène de dimorphisme sexuel.
La saga continue avec un nouvel article publié en 2022 dans Nature où un fémur, découvert il y a plus de 20 ans, vient d’être officiellement décrit et dont l’étude montre qu’une douzaine de caractères démontre la capacité à la bipédie de S. tchadensis bien que celui-ci grimpait encore très certainement aux arbres. Enfin, en 2023, Marc R. Meyer et al. publient une étude qui soulève l’hypothèse d’une locomotion de type knuckle-walking ! Ceci exclurait S. tchadensis des Hominines.
Vous l’aurez compris, bien que toujours considéré comme faisant partie des potentiels ancêtres de la lignée humaine, la position de S. tchadensis est extrêmement débattue.
Orrorin tugenensis
L’espèce Orrorin tugenensis est créée en 2000 suite à la découverte d’une douzaine de restes dentaires et postcrâniens à Tugens Hill au Kenya par Martin Pickford et Brigitte Senut. Ces restes sont datés entre 6,1 et 5,7 Ma.
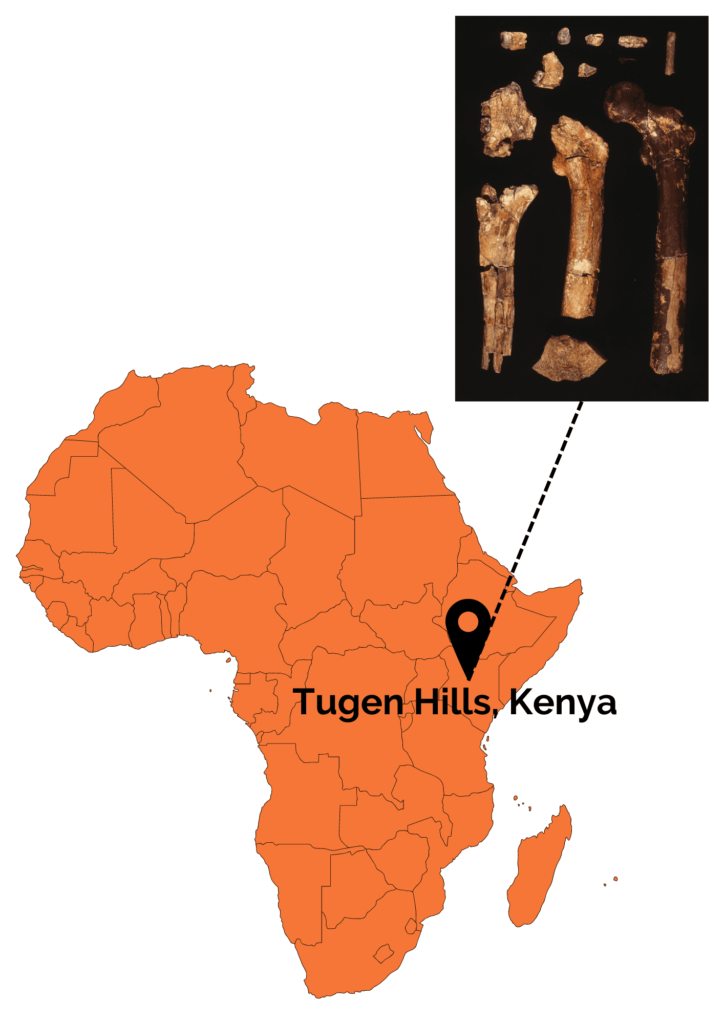
Plusieurs caractères morphologiques rapprochent O. tugenensis des Hominines plus récents. Au niveau de la dentition, les molaires retrouvées sont de forme carrée et de petite taille en comparaison avec les autres primates non-humains. De plus, les canines sont de taille réduite et il y a une absence de diastème. A l’inverse des chimpanzés, les molaires supérieures d’Orrorin présentent des degrés d’usure différents, ce qui suggère une croissance différentielle des molaires. En effet, la première molaire, qui est la plus usée, serait sortie avant la seconde molaire puis enfin la troisième molaire qui n’est presque pas usée. Cette croissance différenciée des molaires se retrouvent bien sûr chez H. sapiens mais également chez les Australopithèques.
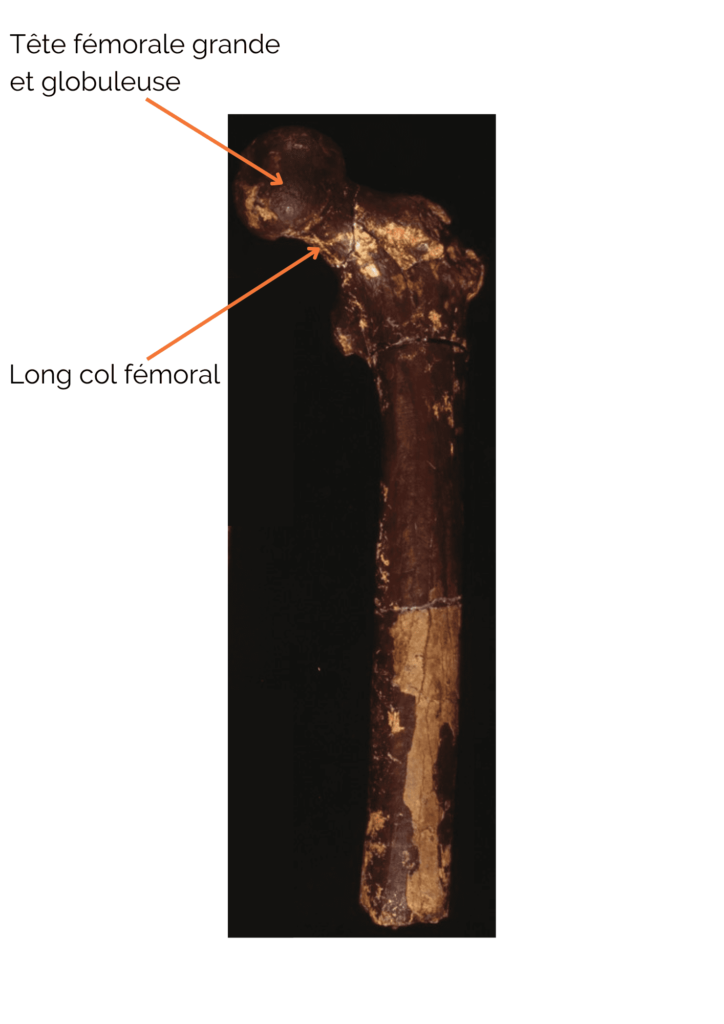
Les restes postcrâniens montrent plusieurs indices allant dans le sens d’une locomotion bipède régulière. Parmi ceux-ci, nous pouvons citer un long col fémoral avec une tête globuleuse et grande, se rapprochant de la disposition observée chez Homo sapiens. De plus, l’os cortical au niveau du col fémoral présente une épaisseur importante. Ceci est un bon indice de bipédie car cela est interprété, chez les bipèdes, comme un renforcement afin de soutenir le poids du tronc sans risque de cassure. Néanmoins, bien qu’Orrorin ait probablement été bipède occasionnel ou régulier, sa bipédie ne ressemblait probablement pas à la nôtre. Il faut également souligner que les os des membres supérieurs, notamment une phalange de la main dont la forme est incurvée, montrent quant à eux des adaptations au grimpé dans les arbres. Orrorin était donc à la fois capable d’arboricolie et de bipédie ! Au niveau de son environnement, les études ont montré qu’Orrorin évoluait dans un environnement de type forêt tempérée sèche avec la présence de zones humides. Ceci a été très discuté au sein de la communauté scientifique car cela entre en contradiction avec la théorie de l’émergence de la bipédie par redressement du corps dû à un environnement ouvert. Les études des dents suggèrent qu’il était frugivore ou omnivore.
Le genre Ardipithecus
Le dernier candidat au titre de plus ancien Hominine est le genre Ardipithecus. Deux espèces appartiennent à ce genre :
- Ardipithecus kadabba qui est daté entre 5,77 et 5,2 Ma et dont les fossiles ont été découverts dans différentes localités de la moyenne vallée de l’Awash en Ethiopie
- Ardipithecus ramidus qui a également été découvert dans la moyenne vallée de l’Awash en Ethiopie et qui est daté aux alentours de 4,4 Ma
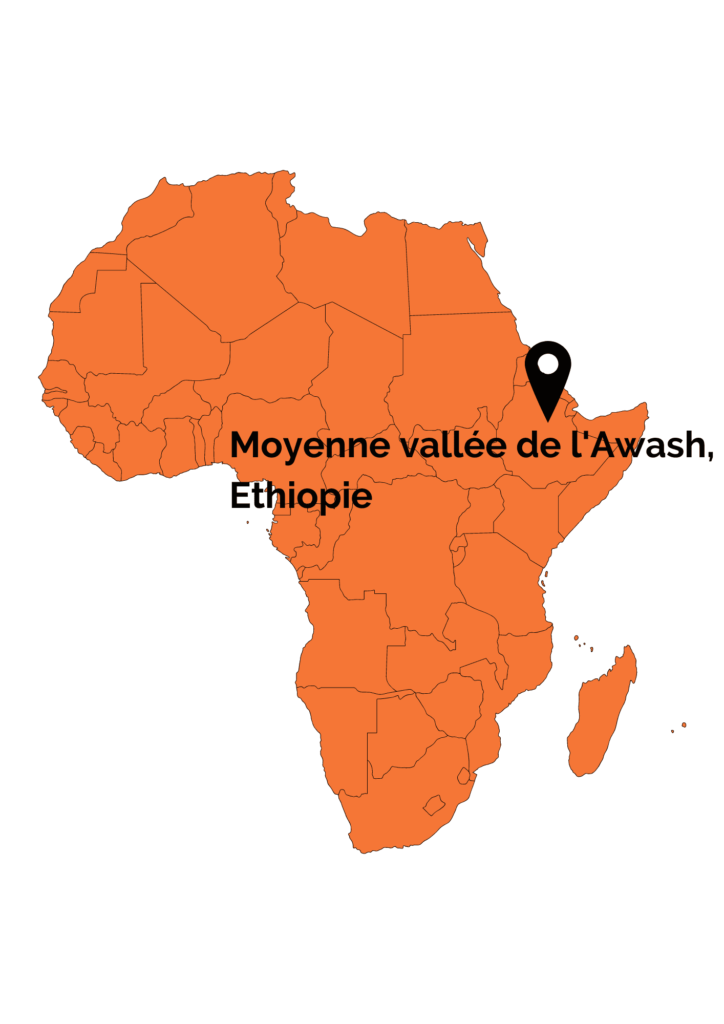
A. kadabba présente de nombreux caractères primitifs comme par exemple des canines de grande taille impliquant la présence d’un diastème ainsi qu’une articulation du coude particulièrement solide indiquant une locomotion par brachiation. De même, les phalanges des mains et des pieds sont longues et courbes, ce qui indique une capacité à l’arboricolie. Quelques caractères rapprochent néanmoins cette espèce de la lignée humaine. C’est le cas par exemple de la surface articulaire d’une phalange proximale de pied qui présente une inclinaison dorsale. Ce caractère est considéré comme étant unique aux bipèdes, ce qui relierait ainsi A. kadabba aux Hominines plus récents. A. kadabba est parfois vu comme étant l’ancêtre de la seconde espèce connu appartenant au genre Ardipithecus : Ardipithecus ramidus.
Tout comme son potentiel ancêtre, Ardipithecus ramidus présente également des caractères indiquant à la fois une locomotion arboricole et une capacité à une locomotion bipède.
Bien que la place de ces 3 genres au sein des Hominines reste débattue, leur importance afin de comprendre l’origine de la lignée humaine est quant à elle certaine ! En effet, ces spécimens se situent à une période où nos ancêtres divergent des ancêtres des chimpanzés, nos plus proches cousins, et illustrent ainsi le début de l’histoire humaine.
Nous remercions Amélie Vialet, maître de conférence spécialisé en paléoanthropologie, d’avoir relu la première version de cet article.
N’hésitez pas à nous poser vos questions et à nous faire part de vos remarques sur le blog. Vous pouvez aussi nous contacter par e-mail. Retrouvez-nous également sur Instagram, Facebook, Twitter, TikTok, LinkedIn ainsi que YouTube pour suivre toutes nos actualités !
A très bientôt,
L’équipe de Prehistory Travel.
Bibliographie :
[1] Michel Brunet, « Sahelanthropus tchadensis dit « Toumaï » : le plus ancien membre connu de notre tribu », Bulletin de l’Académie Nationale de Médecine, 204 (2020)
[2] Michel Brunet et al., « Toumaï, Miocène supérieur du Tchad, le nouveau doyen du rameau humain », Comptes Rendus Palevol, 3 (2004)
[3] G. Daver et al., “Postcranial evidence of late Miocene hominin bipedalism in Chad”, Nature, 609 (2022), https://doi.org/10.1038/s41586-022-04901-z
[4] Martin R. Meyer et al., “Knuckle-walking in Sahelanthropus? Locomotor inferences from the ulnae of fossil hominins and other hominoids”, Journal of Human Evolution, 179 (2023), 103355
[5] M. Pickford et al., “Bipedalism in Orrorin tugenensis revealed by its femora”, Compte rendus Palevol, 1 (2002)
[6] Y. Sawada, “The age of Orrorin tugenensis, an early hominid from the Tugen Hills, Kenya”, Comptes rendu Palevol, 1 (2002)
[7] B. Senut, « Orrorin tugenensis et les origines de l’homme : une synthèse », Bulletin de l’Académie Nationale de Médecine, 204 (2020)
[8] Peter A. Stamos et al., “Hominin locomotion and evolution in the Late Miocene to Late Pliocene”, Journal of Human Evolution, 178 (2023), 103332
[9] Tim D. White et al., “Ardipithecus ramidus and the Paleobiology of early hominids”, Science, 326 (2009)


